Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
4.5.2 Positional Assembly Using Proteins
Bradbury [2045] proposes using artificial enzymes for nanoscale parts assembly. After noting Merkle’s [2017] suggestion that the positional control of nanoscale building blocks would allow the extension of the normal reactions found in biological systems to include free-radical chemistry and more recent descriptions of similar reactive chemistries being employed by enzymes [2046], and related informal suggestions for artificial mechanoenzymes by Freitas [2047] and “mechalysts” by Craver [2048], Bradbury [2045] envisioned the engineering of artificial multifunctional proteins called “robozymes” having the following properties: (1) unfolded, the robozyme grabs onto molecular building blocks [2088-2090], carefully keeping them separate from each other to avoid nonspecific reactions; (2) using specific enzyme catalytic sites near the bound building blocks, it “activates” the molecules (perhaps producing one or more free radicals); (3) induced folding brings the building blocks into relatively precise alignment allowing the desired chemical reaction(s) to occur; and (4) the protein is induced to unfold, releasing the final product. With protein folding forces in the tens to hundreds of pN, such enzymes could also provide a means for threading one molecule through another molecule, mechanically producing interlinked structures such as rotaxane and catenane [1516] without using self-assembly (i.e., by adding hydrophobic ring structures to the molecular parts which will then be attracted to each other in a polar solvent like water) and thus allowing the positional assembly of very small (<1 kD) molecular nanoscale parts.
Note that while most enzymes in cells are involved in manipulating small molecules <0.25 kD, there are several classes of enzymes involved in manufacturing complex covalently bound molecules such as vitamins, enzyme cofactors, antibiotics, and toxins with masses up to ~3 kD. Molecules even larger than this are manipulated by tRNA-synthetase (a 40-100 kD enzyme that manipulates ~30 kD tRNAs), the spliceosome, the ribosome, the proteosome, and the DNA replication complex. (Many of these also involve “parts insertion” or “threading” maneuvers, such as the clamp and bridge helix mechanisms in RNA polymerase II that act as a translocation ratchet to feed DNA through the enzyme interior in order to produce mRNA [2049].) By designing synthetic enzymes consisting of synthetic amino acids, we can envision grabbing molecular parts in a solution and then, as the enzyme folds, bringing them into proper alignment and causing them to react – which might be called “nanopart synthetases” or “protein-directed parts assembly.” Of course, RNA-based ribozymes [2050-2054] may prove better suited than proteins for some reactions, so we are not limited to using enzymes to form the covalent bonds required in nanoparts. Directed evolution in bacteria might be employed to create such specialty enzymes – as has already been done, for example, in converting a disulfide reductase (disassembly) enzyme into a disulfide-forming (assembly) enzyme [2055].
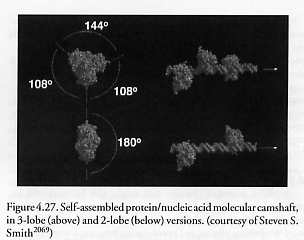
Ratchet-action protein-based molecular motors are well-known in biology [2056]. Conformational cascades of a special genetic variant of yeast cell prions have already been used to self-assemble silver- and gold-particle-based nanowires [2057, 2058]. The GTPase dynamin mechanoenzyme – which self-assembles into rings or spirals, wrapping around the necks of budding vesicles and squeezing, pinching them off, during cellular endocytosis – is well-known [2059-2063]. Amyloid fibril lamination can be exploited for nanotube self-assembly [2064], and artificial self-assembled peptides can orient carbon nanotubes (CNTs) into fibers [2065], achieve hydroxyapatite crystallization [2066, 2067], or grow copper nanocrystals [2068]. Smith [1153, 2069] has used methyltransferase-directed addressing of fusion proteins to DNA scaffolds to construct a molecular camshaft (Figure 4.27) as a exemplar protein/nucleic acid biostructure. Bachand and Montemagno [2070] have engineered a biomolecular motor constructed of F1-ATPase protein [2071] with an attached silicon nitride “propeller” arm [2072] and a reversible on/off switch [2073], and other task-optimized genetically engineered molecular motors have been synthesized by others [2074]. Protein-protein binding specificity has been used to bend silicon microcantilevers [2002, 2004, 2007]. Finally, molecular chaperones are a group of proteins that assists in the folding of newly synthesized proteins or in the refolding of denatured proteins. Genetically engineered chaperonin protein templates (heat shock proteins serving as chaperone molecules) can direct the assembly of gold (1.4, 5, or 10 nm) and CdSe semiconductor quantum dots (4.5 nm) into nanoscale arrays [2075].
Immunoglobulin (Ig) or antibody molecules could be used first to recognize and bind to specific faces of crystalline nanoparts, then as handles to allow attachment of the parts into arrays at known positions, or into even more complex assemblies. As reported by Freitas [235] (next 3 paragraphs): Kessler et al [2076] raised monoclonal antibodies (MAbs) specific for crystals of 1,4-dinitrobenzene having well-defined molecular-level structures. These antibodies were so specific they would not bind to the same molecule when it was conjugated to a protein carrier. IgG antibodies isolated from the serum of rabbits injected with crystals of monosodium urate monohydrate or magnesium urate octahydrate evidently bear in their binding sites an imprint of the crystal surface structure because they can act as nucleating templates for crystal formation in vitro with extremely low cross-reactivity, despite the similar molecular and structural characteristics of the two crystals [2077]. Antibody binding to monosodium urate crystals has been known for decades [2078], and viruses have been engineered with a specific recognition moiety for ZnS nanocrystals used as quantum dots [2160]. Interestingly, antibodies specific to in vivo water-ice crystals have even been reported [2079].
Like antigens with ordered multiple epitopes (the antigenic determinants), crystals expose chemically and geometrically distinct surfaces, so different antibodies might recognize distinct faces of a crystal (possibly including diamond or sapphire crystal faces) in an interaction similar to that of antibodies for repetitive epitopes present on protein surfaces [2080, 2090]. For instance, one MAb to 1,4-dinitrobenzene crystals was shown to specifically interact with the molecularly flat, aromatic, and polar (101) face of these crystals, but not with other faces of the same crystal [2088]. MAbs have also been elicited against cholesterol monohydrate crystals [2081, 2082], one of which [2081] was shown to specifically recognize the crystal’s stepped (301) face. Here, the hydrophobic cholesterol hydrocarbon backbone is exposed on one side of the molecular steps while hydroxyl residues and water molecules are exclusively exposed on the other side. In both cases, crystal-specific antibodies were of the IgM idiotype [2090]. This accords with the assumption that (unlike most commonly used antigens) crystals cannot be processed by the antigen presenting cells, hence antibodies must be induced through a T cell-independent path [2083]. Semiconductor-binding [2160, 2163] and calcite-binding [2084] proteins are known that can discriminate among the various crystal faces of the given material and can in some cases alter the pattern of crystal growth [2085]. Sulfur-free gold-binding proteins (GBPs) recognize and noncovalently bind preferentially to the Au (111) crystal surface – GBPs use multiple repeats of 14-30 residue sequences to bind to this surface [2086]. Hyaluronan is believed to be a crystal-binding protein for calcium oxalate monohydrate crystals [2087].
Solubilized (derivatized) C60 and C70 fullerenes can induce the production of specific antibodies [2089-2093], usually by interaction with the combining sites of IgG [2089]. It is speculated that highly hydrophobic pure fullerenes would be recognized by antibodies with hydrophobic amino acids in their binding sites [2089, 2094] or would interact with donor -NH2 [2098] and -SH [2099] groups. There are now many reports of antibodies being raised to single-walled carbon nanotubes [2093-2097]. For example, antibodies raised to C60 in mice strongly bind to single-walled nanotubes [2097]. Computer simulations suggest that it may be possible to build antibodies that selectively bind to nanotubes of a specific diameter or chirality [2094].
Last updated on 1 August 2005
{kind=link}