Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
5.1.9.L Evolvability
Note that evolvability [2568-2570] (see also Sections 2.1.5 and 5.2) is an undesirable characteristic for artificial replicating systems intended to serve as molecular manufacturing systems (Section 5.11). An important key to suppressing evolvability and the “evolution of diversity” [2571] is to suppress variability – that is, to avoid multigendered systems and to ensure strict replication fidelity. A digital asexual reproductive system, one that only produces the same design generation after generation, can be employed to avoid evolvability. Evolvability must be actively avoided because evolvable hardware, if allowed to progress unchecked, is likely to develop capabilities faster than “centrally-planned machinery,” a temptation that runs counter to our desire for strong public safety (Section 5.11).
Smith [2572], following Eigen and Schuster [1663], defined three criteria sufficient to consider replicators as “units of evolution”: (1) multiplication (replicators give rise to more of the same kind); (2) heredity (like entities produce like offspring); and (3) variability (inexact heredity). Objects of different types having a hereditary difference in fecundity or survival produce evolution in the replicator population via natural selection. But without variability, the replicators are merely “units of selection”, and evolution by natural selection cannot occur. Still, we must be cautious. C. Phoenix [572] notes that “even a limited feature set is still susceptible to what one might call physical mutation. For example, a construction error in a robot arm might cause arms that it builds to be more likely to have the same error, even with an unchanged blueprint.” Similarly, Yudkowsky [2452] asks: “An important question is whether the replicator design allows for the equivalent of ‘prion’ disorders – that is, a ‘self-replicating’ protein misfold that propagates without the need for mutated DNA. For example, suppose that a certain manufacturing defect within a lever arm does not break the lever arm, but reliably results in the same defect being present in any lever arms manufactured with the assistance of the first lever arm. There would then be heritable information between replicators of a type that bypasses the manufacturing instruction streams.”
More broadly, Yudkowsky [2452] notes that whether a given design space is evolvable may depend on the probability distribution for mutations, and whether there exists any probable mutation that leaps to a different viable point in the design space: “Has the replicator design been audited to ensure nonevolvability relative to a possible space of probable point mutations?”
L1. Gendericity. The implications
of gender in artificial self-replicating machines have only been lightly addressed,
even in the realm of cellular automata [2573].
As Merkle [2574] noted in 1989: “In
nature, self-replicating systems evolve. A major mechanism supporting this evolution
is sex. Even bacteria have sex of a sort (they exchange DNA quite promiscuously).
This is a critical component in the ability to evolve. I would therefore suggest
that we not include sex in artificially designed self-replicating systems. If
there are legitimate research needs for exploring these options, they should
be done under tight constraints and should be approached with a great deal of
caution.” Vitanyi [2573] suggests
that a transition from asexual to sexual reproduction is necessary to produce
a change in the number and structure of the genetic tapes employed by an organism.
A variety of interesting replication-control mechanisms for gendered replicators
have been (instructively) explored by nature, as for instance the abortive karyogamy
employed by Wolbachia [2575],
an endosymbiont rickettsial bacterial species thought to infect the reproductive
tissues of up to 20% of all insect species [2576].
L2. Offspring Fertility.
Can replicator offspring (i.e., daughter devices) produce further copies of
themselves? The “fertility test.”
L3. Number of Fertile Mutant Configurations
(aka. “heredity”). Szathmary [2416]
notes that autocatalytic replication in the formose system [2419]
“is entirely non-informational, there are no hereditary variants.”
On the other hand, in animal breeding there are a vast number of fertile configurations,
allowing breeders to explore a large contiguous phenotypic space – for
example, as Mark Tilden notes: “The dog is all the things you can do with
a wolf.” (Anecdotal evidence claims an increasing incidence of natural
births of elephants lacking tusks [2577]
and (less likely) of rattlesnakes lacking rattles [2578],
in response to human predation, and the acquired resistance of bacteria to antibiotics
is well-known [2579], though species already
at their environmental limits may not be able to adapt to further change [2580].)
In self-replicating robots, a checksum function embedded in the replication program could affirmatively prevent replication of mutant configurations [2581]. (In effect, the checksum serves as an activation key: if the key is incorrect all further construction is halted, or else the system tags itself for disposal or even self-destructs.) An example of active replication data self-policing is found in nature: Tetrahymena, a single-celled eukaryote, deletes up to 15% of its somatic nuclear DNA during each replication cycle in a process called “programmed DNA deletion” that can recognize and delete foreign genes that have invaded a chromosome [2582]. This property of replicators is also known as brittleness or “canalization” [2401, 2583]. It has recently been proposed that an electronic self-test for base pair mutations may occur naturally in DNA that is functionally equivalent to checksum correction [2584].
From a pragmatic engineering perspective, it is desirable that small shifts within the design space reliably result in nonviable designs. Notes Yudkowsky [2452]: “All evolved organisms are very highly evolvable by virtue of having evolved. A consciously executed human design tends to leap large areas of design space and contain precise patterns that break easily. Even when minor (probable, point) shifts in the design permit a functionally similar result, the large-scale architecture may still be fragile enough to constrain the design to a particular volume of the design space.” Natural biological replicators and artificial replicators designed using evolutionary algorithms are the result of a design process that optimizes by climbing along an incremental pathway composed of exactly the kind of probable point mutations that would be likely to occur in a replicator design that we would prefer to hold constant. Artificial replicators designed using nonevolutionary algorithms are produced by a series of cognitive manipulations that differ fundamentally from the types of point mutations that would be produced by random physical errors, and random physically induced errors in these machines would be unlikely to leap to viable points, let alone to superior points, in the design space.
L4. Replication Fidelity.
How faithful is the copy to the original? Luksha [2388]
considers the similarity between a producer and what is produced to be a key
property of self-reproduction as a general phenomenon. He refers to this dimension
as the “parent-progeny relationship” and suggests three classes
of self-reproduction including “exact replication,” “near
replication of an ancestor,” and “near replication of a parent.”
This design factor may be captured by the following unidimensional vector:
In a personal correspondence, Luksha [128] clarifies this analysis by using a two-dimensional grid in which the horizontal axis represents replication fidelity relative to the original replicator and the vertical axis represents replication fidelity to the replicator’s immediate ancestor. To quantify fidelity, Luksha introduces a qualitative difference function d(a,b) between an earlier replicator system a and a descendent replicator system b, which is a measure of the distance between the two points a and b in a space of relevant replicator characteristics (e.g., using two such characteristics x and y, then d(a,b) = {(xa – xb)2 + (ya – yb)2}1/2). Luksha then considers replicator system st produced by its immediate ancestor st-1 in generation t, and also some initial system s0 which starts the reproduction of the given type of systems, where distance D is a level of acceptable variation (e.g., see Eigen et al [2585] for similar measures of this kind used in pre-biotic models). This yields the following two-dimensional classification scheme:
Two cases are only theoretically possible. In case [A], the system has evolved from the original state to some “attractor” state which is reproduced “exactly”. “Theoretically, this is a possible case; however, its mechanics are not clear if non-trivial self-replication is considered: a system must conserve its copying to a given structure, but since it has evolved to this structure it has potential for further evolution.” Case [B] is another theoretically possible situation where “a system which has evolutionarily ‘departed’ from a given system occasionally returns to this system.”
The effects of copy error on the time evolution [2586] and survival [2587] of simple self-replicating systems have been analyzed by others. Note also that the exact physical duplication at the quantum level is evidently impossible [276].
L5. Genomic Redundancy.
Greater genomic redundancy is believed to contribute to increased evolvability
of a replicator [2588] – “populations
of robots with larger genotypes achieve systematically higher fitness than populations
whose genotypes are smaller.” Note that this will not be a consideration
in regard to replicators with external non-replicated genomes, such as broadcast-architecture
replicators.
L6. Environmental Partitioning.
Computational studies by Birol et al [2589]
show that partitioning of the replicator-accessible environment into two or
more homogeneous subenvironments “leads to emergence phenomena exemplified
by steady states not obtainable in the equivalent homogeneous system. The coupled
environments can host species that would not survive should the coupling be
removed. Two coupled environments may have regions in parameter space that result
in chaotic behavior, hence segregation in the environment causes complexity
in the system dynamics.” Hence environmental partitioning may result in
greater evolvability (among those classes of replicators capable of mutation),
and thus should be avoided when designing stable manufacturing systems. For
replicators inherently incapable of mutation, environmental partitioning is
not of concern.
L7. Complexification. “Over
the course of biological evolution, new genes are occasionally added to genomes,
increasing the complexity of the phenotype. Complexification has led to major
innovations in body-plan organization.” [2401]
Implementing a mechanism for handling variable length genomes would enable replicator
complexification, thus enhancing evolvability in artificial replicators. This
should be avoided in “safe replicator” designs. Complexification
also is more likely to occur if information caches, parts, or subunits serve
multiple unrelated functions (e.g., B12, E9,
F7).
L8. Heterochrony. Changes
in the timing or ordering of developmental events over generations of replicators
is called heterochrony [2401, 2590].
In natural (biological) embryogeny, the path to the final product is surprisingly
flexible – entire phases of development can be eliminated without sacrificing
the end result. “For example, many frog species have evolved away their
tadpole stage, yet still grow into mature, sexually functioning frogs [2591].
Their limb buds develop early, and a little frog, rather than a tadpole, ultimately
emerges from the egg. Contrariwise, the Mexican axolotl, a salamander, has lost
its adult stage, and develops mature gonads as a tadpolelike creature
[2592]. Their development of gonads has
been greatly accelerated so that they become sexually functional as tadpoles.
This dramatic flexibility in development suggests that significant modularity
underlies the genetic encoding. Because the timing of the development of different
modules is so flexible, mutations can safely modify timing, allowing evolution
to explore a variety of developmental plans. Heterochrony allows developing
components to come into contact with different components, so that evolution
can explore many points of synthesis between components....In addition, heterochrony
increases the number of successful genotypes by offering a variety of paths
to each successful phenotype.... Like Cartesian coordinates being used in lieu
of chemical gradients, time itself can be used to regulate genes. Since time
is directly available, it can be exploited as a ‘growth hormone’
that explicitly activates and terminates events in embryogeny.” [2401]
Similarly, the JohnnyVon replicator simulation (Section
2.2.2) includes a single entity that switches roles over time, changing
from genotypic replication in its youth to phenotypic production in its maturity.
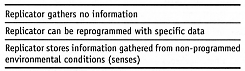
L9. Programmability and Learning.
Can the replicator respond to, or learn from, the environment, a key component
to evolvability? The incorporation of information storage and feedback mechanisms
in supramolecular prebiotic assemblies has been called “crossing the Darwinian
threshold,” a key step in the origin of life [1726].
A replicator that can be reprogrammed is a simple example of learning from the
environment. Safety is reduced when replicators are increasingly able to gather
information from more diverse sources in the environment.
L10. Mode of Genomic Heritability.
The manner in which self-descriptive information is passed on to offspring determines
the modes of evolution that are possible. How, and in what manner, can replicators
pass along random-access information to progeny? Safety is reduced when replicators
can pass along more versatile memory stores to their offspring.
L11. Multilevel Genomic Heritability.
Is genomic information heritable only from the level of the whole replicator,
or can individual subunits within the replicator pass on heritable information
independently of the whole? In regard to metazoan evolution, Hoenigsberg [2593]
notes: “The distinction between heritability at the level of the cell
lineage and at the level of the individual is crucial. While all out reproduction
is the Darwinian measure of success among unicellular organisms, a high replication
rate of cell lineages within the organism may be deleterious to the individual
as a functional unit. If a harmoniously functioning unit is to evolve, mechanisms
must have evolved whereby variants that increase their own replication rate
by failing to accept their own somatic duties are controlled. Modifiers of conflict
that control cell lineages with conflicting genes and new mutant replication
rates that deviate from their somatic duties had to evolve. The metazoan embryo
is not immune to this conflict especially with the evolution of set-aside cells
and other modes of self-policing modifiers [2594].
In fact, the conflict between the two selection processes permitted a Lamarckian
soma-to-germline feedback loop. This new element in metazoan ontogeny became
the evolvability of the vertebrate adaptive immune system and life as we know
it now. We offer the hypothesis that metazoan evolution solved this ancient
conflict by evolving an immunogenetic mechanism that responds with rapid Lamarckian
efficiency by retaining the ancient reverse transcriptase enzyme (RNA©DNA)
copying discovered by Temin [2595] in 1959
and found in 1970 in RNA tumor viruses by Temin [2596]
and Baltimore [2597], which can produce
cDNA from the genome of an RNA virus that infects the cells.” Cell lineage
selection within the individual replicator can alter the rate and fixation probability
of new mutations [2598].
L12. Interreplicator Information
Transfer. Can replicators of different kinds exchange genomic information?
In biology, horizontal gene transfer [2599]
is the collective name for processes that permit the exchange of DNA among organisms
of different species. Replicator safety is enhanced if replicators of different
kinds lack the ability to exchange genomic information among themselves.
Last updated on 1 August 2005