Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
5.2 Replication Time vs. Replicator Mass
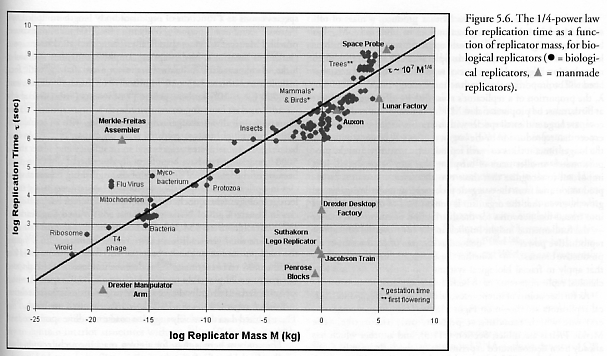
Figure 5.6 shows the replication time (t) as a function of replicator mass (M) for 126 biological species and 9 actual or proposed artificial kinematic self-replicating systems across a size range spanning nearly 20 orders of magnitude, using data drawn from numerous sources [2600] and tabulated in Appendix A. (The exact formula for the trendline drawn in Figure 5.6 is t = 1.78 x 107 M1/4.) It is well-known [2601] that the time of fecundity, the age for maturity, and the regeneration time grow with replicator size. For example, as long ago as 1965 Bonner [2602] found a strong nearly-linear positive correlation between body length and the logarithm of generation time. Our data confirm that replication time in biological systems appears to follow a 1/4-power law function of replicator mass, as explained by the model for allometric scaling laws in biology recently proposed by West et al [2540]. Indeed, age at first reproduction can be shown from first principles [2603] to be proportional to M1/4, a value which has elsewhere been substantiated empirically [2604, 2605]. On the other hand, it is also known that nonfractal mechanical systems such as combustion engines and electric motors exhibit purely geometric 1/3-power, rather than 1/4-power, scaling with mass [2608-2610]. It has been expected that for a given replicator architecture, physical replication speed should be a linear function of geometric dimension (hence a 1/3 power of mass, since mass is volumetric) because nanomanipulator velocity is scale invariant [208] while travel distance decreases linearly with dimension. (See also Section 5.9.5.)
Figure 5.6 may provide one simple answer to a question that is sometimes asked in relation to machine self-replicating systems (SRS) [988]: “If self-replication is so easy, where are all the SRSs?” The chart suggests that pre-nanotechnology macroscale replicators may be expected to have comparatively lengthy replication times. For example, the replicating space probe proposed by Freitas (Ref. [1014], Section 3.11) was scaled at 4.43 x 105 kg (443 tons) dry mass and a replication time of 1.69 x 109 sec (53.7 yrs), the replicating lunar factory proposed by NASA in the early 1980s (Freitas and Gilbreath [2], Section 3.13.2) was scaled at a dry mass of 105 kg (100 tons) and a replication time of 3.14 x 107 sec (1 yr), and the Lackner-Wendt auxons (Section 3.15) were scaled to a ~20 kg mass and ~1.2 x 107 sec replication time [1128]. All lie close to the M1/4 trendline and suggest that autonomous meter-sized (or larger) replicators that do not make use of nanoscale manufacturing technologies may have replication times on the order of months or years. Solid printers (Section 3.20) typically weigh 200-500 kg with an effective “replication” time of 1-3 months (about 1 order of magnitude below trendline). The replication time of a typical modern industrial park or machine shop may also be on the order of months to a year, so to obtain a millionfold expansion of factory mass one must wait ~20 replication cycles (i.e., 220 ~ 106), or ~2-20 years. The conventional economics of discounted present value, plant depreciation, and 3-5 year venture capital investment horizons would render most such projects commercially nonviable. Investing today in a seed device that one must pay skilled people to teleoperate for 2-20 years during the non-productive growth phase, yielding at the end of that time a 2-20 year old, possibly nearly obsolete, factory makes little economic sense.
However, using nanotechnology-based replicators having replication times on the order of 1000 seconds allows a millionfold capacity increase in only 200,000 sec (~2.5 days) – that is, rapid prototyping, fast factory buildout, and quick movement to market with useful products made in a nonobsolete manufacturing facility. The scaling laws for replication speed as a function of replicator size thus appear to interact with the economic and obsolescence factors to produce a viable scenario for the deployment of commercial replicators only in the microscale, but not in the macroscale, realm.
Because biological replication time scales as M1/4, larger replicators must spend a longer absolute time accumulating resources and maturing tissues than do smaller replicators [2611]. Additionally, in biology a smaller individual replicator produces a mass of offspring that represents a larger fraction of its adult mass than a larger replicator [2611], and smaller animals generally produce more offspring [2558] – further facts which argue for the comparatively greater commercial value of microscale manufacturing systems that offer rapid ramp-up of output capacity and higher production per unit mass of factory machinery.
Consider a replicator of mass M that produces a mass of offspring
Moff = lM that is some fraction l
of its adult mass. M
As further points of interest, several additional artificial mechanical replicators are shown on Figure 5.6 – including (among others) one which remains, at present, only theoretical (the Merkle-Freitas assembler; Section 4.11.3), and another which has already been demonstrated experimentally (the Suthakorn LEGO® replicator; Section 3.23). By the bio-centric measure of this data, the Merkle-Freitas assembler appears very inefficient and seems to require at least 3 orders of magnitude potential improvement in replication speed to catch up with biological systems. Even with this handicap, the device still replicates at least an order of magnitude faster than conventional macroscale industrial machinery. But replication time is not merely a function of mass, as indicated by the seemingly anomalously rapid replication time achieved by the comparatively simple Suthakorn-Chirikjian LEGO® replicator. Replication time is also a function of the complexity of the device, the replicator parts count or parts size, the degree to which the replicator must fabricate components from a disordered feedstock, the degree to which control and processing mechanisms can be offloaded from the replicator device to external systems, and potentially dozens of other dimensions of the replicator design space (Section 5.1.9). It is possible, even likely, that nanotechnology-based replicators can be at least 1-2 orders of magnitude more efficient than biological replicators, size for size, albeit at the cost of higher power density (Nanomedicine [228], Section 6.5). But until comprehensive further studies can elucidate the precise scaling laws governing the replication time of mechanical replicators, natural biological systems may stand as useful benchmarks of the minimum performance that may be expected from future artificial self-replicating systems.
Finally, note also that evolvability scales inversely with size, all else equal. In the first place, in biology the replicator abundance increases as a function of decreasing replicator size [2618-2623], with the number of replicators N(m) as a function of replicator mass m given by N(m) ~ constant · m-2, a Zipf power law distribution believed due to predation-related deaths in macroscale animals [2624]. In the second place, the number of distinct (biological) replicator species is a declining function of the characteristic size of individual species members – i.e., smaller creatures produce more species [2606-2608], as illustrated by the frequency distribution of both insect and vertebrate species count as a function of organism body length compiled by Pianka [2625] and as a function of body mass for 2104 mammals compiled by Maurer [2611]. More specifically: There are ~105 known species [2626] of mammals, birds, reptiles, amphibians, fishes and mollusks, of characteristic individual organism mass ~0.1 kg, with, for instance, an estimated ~3 x 1011 birds alive on Earth [2627]. There are 751,000 (i.e., ~106) known species [2626] of insects (estimated up to ~3 x 107 species counting currently unknown insect species [2628]) of characteristic individual organism mass ~10-5 kg, with an estimated ~1019 individual insect organisms alive on Earth [2629-2631]. As for bacteria, microbial systematics is still in its infancy and fewer than 5000 bacterial species have been formally described [2626, 2632], in part because of the traditional difficulty of distinguishing bacterial species in optical microscopes. But modern measurements using molecular biology techniques have found ~4000 different bacterial species in a mere 1 gm of Norwegian forest soil [2633] and Sugawara et al [2634] have identified 334,312 distinct strains of bacteria that are already known. Even traditional conservative estimates [2635-2637] put the total number of unknown bacterial and fungal species at ~107 but the most recent estimate [2638] is that the unknown bacterial species count could plausibly reach as high as ~108-1013 species, with an estimated ~1031 individual bacterial organisms of characteristic individual organism mass ~10-15 kg currently alive on Earth [2639]. These limited data tentatively suggest a coarse trendline species count of Nspecies(m) ~ 105 m-1/2.
The concern here is that, for a given machine architecture, the smaller the device, the faster that unit reproduction may occur, and hence the greater the number of trial offspring that the device could sire (and test for fitness) per unit time interval. Macroscale reproducers that generate variability only via stochastic genomic mutations are far less likely to be able to generate enough trial offspring to stumble upon successful new phenotypes in any reasonable period of time. Microscale reproducers, on the other hand, may be able to generate offspring up to a million times faster, thus could be far more likely to randomly yield productive modifications, hence to “evolve,” if their design foolishly permits such viable modifications. Microscale replicators may therefore be viewed as inherently more risky from a public safety standpoint [199, 2909] and so the need for adherence to design guidelines [271] to forestall unplanned system evolution is more urgent in this realm.
Last updated on 1 August 2005
{kind=link}