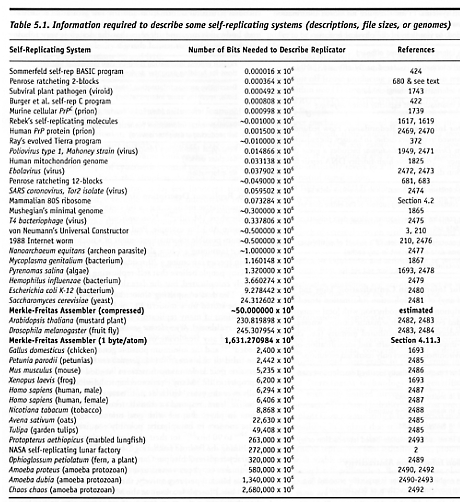
Kinematic Self-Replicating Machines
© 2004 Robert A. Freitas Jr. and Ralph C. Merkle. All Rights Reserved.
Robert A. Freitas Jr., Ralph C. Merkle, Kinematic Self-Replicating Machines, Landes Bioscience, Georgetown, TX, 2004.
5.3 Minimum and Maximum Size of Kinematic Replicators
What is the minimum size for free-standing autonomous kinematic replicators? In microbiology, this question has been tentatively addressed in the past,* with the conclusion that the theoretically smallest biological cells that can be contained within a membrane must have diameters of at least ~50 nm [1774] to 140 nm [1866, 2640]. This question has more recently been analyzed in depth by a special panel of nearly two dozen researchers at a workshop entitled “Size Limits of Very Small Microorganisms,” sponsored by the Space Studies Board of the National Academy of Sciences and first published in 1999 [2641, 2642]. This study was motivated in part by the announcement [2643] of possible evidence for relic biogenic activity in the Martian meteorite ALH84001. This evidence included an observation of “carbonate globules and features resembling terrestrial microorganisms, terrestrial biogenic carbonate structures, or microfossils” that ranged in length from 10-200 nm as revealed in electron microscope images, and also other evidence for nanometer-scale biomineralization and “nanofossils” on Earth [2644-2646] – even though exotic morphological forms resembling biogenic structures can occur via nonbiological processes [2647], and many bacteria are known to undergo transition from a large active state to a small dormant state [2648-2650], often in response to starvation [2648]. The study also was motivated by reports [2653-2660] of “nanobacteria” with calcium shells which have been proposed as possible nucleation sites for kidney stones [2655-2657]. Some of these microbes are claimed to be cultured from blood [2653, 2654] and apparently have diameters as low as 80 nm but average 200 nm (mass ~ 4 x 10-18 kg, replication time ~3 days ~2.6 x 105 sec [2520]). There are also reports of “nannobacteria” as small as 30 nm in diameter present in everything from tapwater to tooth enamel [2661] and “ultramicrobacteria” [2650-2652] with cell volumes of 0.03-0.08 µm3 (386-535 nm spherical diameter) [2651].
* In the 1960s, NASA sponsored research [2662] to inform future astronauts how to recognize even the most rudimentary forms of life on other worlds – with the conclusion that the simplest living thing would contain at least 124 proteins of 400 amino acids each, and would possess a functioning genetic code so the organism would reproduce true to type.
Within the NAS Workshop [2641], geneticists and cell biologists “reached consensus on the smallest size likely to be attained by organisms of modern biochemical complexity. Free-living organisms require a minimum of 250 to 450 proteins along with the genes and ribosomes necessary for their synthesis. A sphere capable of holding this minimal molecular complement would be 250 to 300 nm in diameter, including its bounding membrane. Given the uncertainties inherent in this estimate, the panel agreed that 250 ± 50 nm constitutes a reasonable lower size limit for life as we know it. At this minute size, membranes have sufficient biophysical integrity to contain interior structures without the need for a cell wall, but only if the organism is spherical and has an osmotic pressure not much above that of its environment. Bacteria with a diameter of 300 to 500 nm are common in oligotrophic (nutrient-poor) environments, but smaller cells are not. Nanobacteria (a single-celled microorganism proposed to have a maximum diameter in the range of tens to a few hundreds of nanometers) reported from human and cow blood fall near the lower size limit suggested by cell biologists. However, the much smaller (ca. 50 nm) bodies found in association with these cells may not, themselves, be viable organisms. Observations on Archaea indicate that, in general, they have size limits similar to those for Bacteria. Whereas a cell operating by known molecular rules – with DNA or maybe RNA, ribosomes, protein catalysts, and other conventional cell machinery – would have a lower size limit of 200 to 300 nm in diameter, primitive microorganisms based on a single-polymer system could be as small as a sphere 50 nm in diameter. On Mars or Europa, fossils might preserve a record of biological systems different from those we understand – perhaps early products of evolution that made do with a small complement of functional molecules. Organisms of modern biochemistry might become small by being pathogens or living in consortia – that is, by using the products of another organism’s genes.”
NAS Workshop participant and biochemist Michael Adams [2524] calculated that a 50 nm diameter spherical cell would just have enough room for 2 ribosomes, 520 30-kDa proteins, and 8 1000-bp genes as DNA – well below the minimum possible genome size of ~250 genes [1866, 1867] which would still require the cell to be parasitic, relying on ready-made nutrients from the external environment or a host [2642]. Even Mycoplasma genitalium, the free-living microorganism with the smallest known genome (Table 5.1), has 471 genes [1867] that occupy 10% of cell volume [2641]. So 50 nm is likely too small, concluded Adams [2524]: “...the minimum theoretical size for a cell is 172 nm diameter. To grow, such a cell must be supplied with (and must assimilate) all amino acids, fatty acids, nucleotides, cofactors, etc., because it would contain the minimum number of genes (250) and have a minimal biosynthetic capacity. The [172 nm] cell would have a 5 nm membrane but no cell wall. It would consist, by volume, of 10% DNA, 10% ribosomes, 20% protein, and 50% water, and would contain approximately 65 proteins per gene as well as 65 ribosomes.” By positing a reduced protein size and shifting to single-stranded DNA and segmental genome copying, biophysical chemist Peter Moore [2663] argues that a 60-nm living cell might be possible but must “invoke a biochemistry unlike any known in modern cellular life. A cell that is even smaller would require even more radical departures....their biochemistry will be a lot different from anything we know today.”
For example, Benner [2664] noted that “much of the volume of a bacterial cell is filled with machinery (ribosomes) that convert information in the genetic biopolymer (DNA) into information in the catalytic biopolymer (protein). This places a limit on the size of a two-biopolymer living system that all but certainly excludes cells as small as (for example) the structures observed in the Allan Hills meteorite derived from Mars. Life that uses a single biopolymer to play both genetic and catalytic roles [2665-2671] could conceivably fit within a smaller cell.” Benner suggested that a single-biopolymer system could be packed into a 50-nm sphere. However, he admitted, “no biopolymer has yet been found that can play both roles, and the chemical demands for genetics and catalysis are frequently contradictory.”
A bottom-up biochemical analysis of the minimum requirements for the first primitive lifeform on Earth by Ferris [2672] suggested that if it was possible to have membrane-bounded nonribosomal life with a minimum of five genes (i.e., ligase, replicase, monomer synthase, fatty acid synthase, and membrane synthase ribozymes), then if the organism achieved the single-stranded RNA packing density of Qb virus it would need an interior volume of 3,580-nm3 plus the volume of the surrounding membrane, giving a spherical outside diameter of 27 nm assuming a 4 nm thick membrane. If instead the earliest organism achieved only the double-stranded RNA packing density and protein/biomolecular content typical of contemporary L-A virus, then one would need an interior volume of 48,900-nm3, giving a spherical outside diameter of 53 nm again assuming a 4 nm thick membrane.
Alternatively, NAS Workshop participant Jeffrey Lawrence [2497] hypothesized the possible existence of very tiny self-replicating “cells” based on sequential horizontal transfer of single genes “wandering” among a consortium, illustrating that not all cells need have all the essential genetic information at the same time: “A model is presented whereby the frequency of gene exchange is much greater than the frequency of cell division. In this model, cells may be considered way stations for gene replication and transfer; such organisms need not maintain a full complement of genes, and genome sizes may decrease.” Replication would be very slow (e.g., 10,000 times slower than bacteria, as posited for nanobacteria [2520]) and dependent on the lifetime of biosynthetic intermediates and on the rate of gene transfer, but simulations by Lawrence [2497] predicted “the propagation of organisms where the average cell contains, on average over time, fewer than 1 gene.” Using this strategy, a ~50 nm cell barely large enough to hold 1-2 ribosomes might become feasible. However, another study [2641] participant noted that there could be great selective advantage of aggregating more than one gene into a single compartment by cell-cell fusion, so that even starting with the minimal mechanically stable vesicle size, the system would tend to evolve to larger, more efficient compartments. The greater surface area to volume ratio of smaller cells has been claimed to be adaptive for low-nutrient environments [2673], though counterexamples exist [2674]. In the search for the great universal ancestor of all life, Woese [2675] notes that if there was a great deal of gene transfer, the whole question of the identity of the first cell may become meaningless because the “universal ancestor” of all terrestrial life may not have been one cell, but rather the “genetic annealing” [2675] of a “primordial commune” [2924] of protocells, or as Cowen [2676] describes it, “a kind of cellular mist in which genes transfer from one droplet to the other like molecules of water vapor.” Rasmussen et al [1959] suggest a specific design for a simple protocell capable of self-replicative cycling.
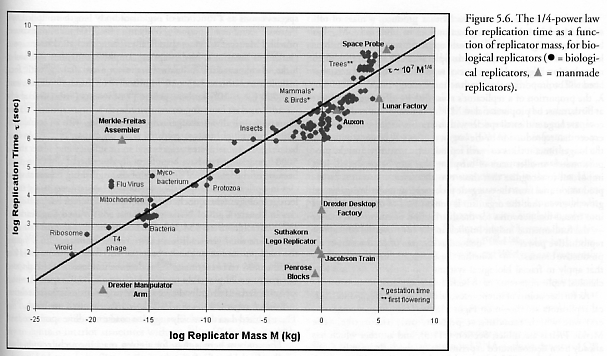
The absolute minimum possible size of purely mechanical (nonbiological) kinematic replicators has not yet been seriously addressed by nanotechnology theorists, but it seems unlikely that a fully autonomous mechanical replicator can be significantly smaller than the ~100 nm size range. However, just as the ~25 nm ribosome is much smaller than the smallest known well-documented free-standing biological cell, it is conceivable that remotely powered and fully-teleoperated nanomechanical manipulators capable of assembling themselves from a handful of prefabricated parts using one or a few simple mechanical processes might be possible at the ~10-20 nm scale. Future research will doubtless clarify this point. Using the replication time/mass relation shown in Figure 5.6, a 20-nm teleoperated replicator with normal density could have t ~ 200 sec.*
* The same time/mass relation would predict a replication time of t = 3.6 sec for a replicator having the limit mass of a single proton, neutron, or hydrogen atom (or t = 0.55 sec for a single-electron-mass replicator) that could somehow replicate solely by the manipulation of covalent bonds and other conventional interatomic forces. But self-replicating entities that employ nuclear forces can replicate much faster and thus are not represented by Figure 5.6. Neutrons are one well-known example of such a nuclear replicator: a single thermal neutron, upon entering a substrate consisting of a 92U235 nucleus, induces fission of that nucleus in ~10-12 sec, replicating 2-3 daughter neutrons which are emitted from the substrate along with various larger fissile fragments, leading to a macroscopic chain reaction if a critical mass of substrate is present [2987].
The maximum possible size of a kinematic replicator is presently unknown, but is likely extraordinarily large. Consider the purely speculative possibility of audaciously extrapolating the known relationship between replication time and replicator mass shown in Figure 5.6 by another 7 orders of magnitude, beyond the current 10 orders of magnitude in time for which we already have extensive real-world replicator data. If the maximum possible replication time approximates the current estimated age of the universe, or t ~ 1.37 x 1010 years (= 4.30 x 1017 sec) [2677], then from the empirically observed relation t ~ 1.78 x 108 M1/4 (Section 5.2) we would compute that the largest possible replicator that could demonstrably exist (i.e., having executed at least one full replicative cycle) would have a mass of M ~ 3.4 x 1041 kg, or about one-third of the mass of our own Milky Way galaxy (~9.8 x 1041 kg, including ~400 billion visible stars and ~4.9 x 1011 solar masses [2678]). Information/materials traveling at/near the speed of light (c = 3 x 108 m/sec) can make NRt ~ c t / Drepl round trips across a replicator of diameter Drepl. If the mass is distributed throughout a galaxy-sized replicator with Drepl ~ 100,000 light-years, then NRt ~ 105. If the mass is concentrated in a replicator approaching maximum possible density (i.e., within the Schwarzschild radius of a nonrotating uncharged black hole), then Drepl ~ 4 GM/c2 ~ 1015 meters (taking gravitational constant G = 6.67 x 10-11 N-m2/kg2) and so NRt ~ 1011. For comparison, a human body with a replication time of t = 0.736 year (~9 months) and a bloodstream circulation time of ~1 minute [228] (using fluidic/chemical information transport at <1 m/sec) has an effective NRt ~ 106.
Last updated on 1 August 2005
{kind=link}
{kind=link}